14) Immunology
I am not aware of any studies.
15) Pathological features
Griner (1983) reported on findings in 72 animals. By that time there had already been 77 births at San Diego , of which 24 had died neonatally and 6 were perinatal deaths. Trauma and a variety of infectious causes of death are listed in that review. Rumenitis with ulcers also occurred and one neonate had a cleft palate. Interestingly, four animals were icteric at birth for which the cause was not identified but possibly linked to a new male father. Baumeister et al. (1983) in a study of rotavirus infection of zoo animals found it primarily in young animals. Appleby et al. (1992) found the trichostrongylid nematode Cooperioides antidorca in intestinal nodules of animals in Kenya .
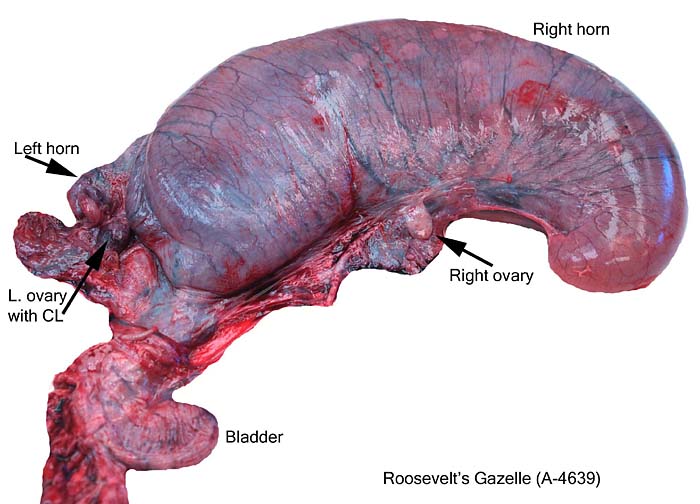
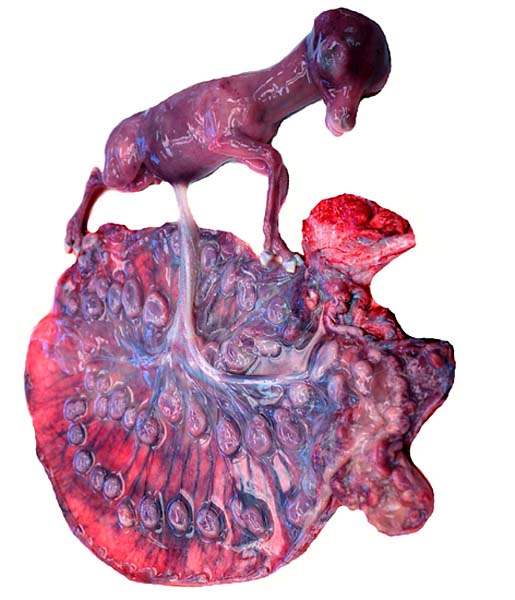
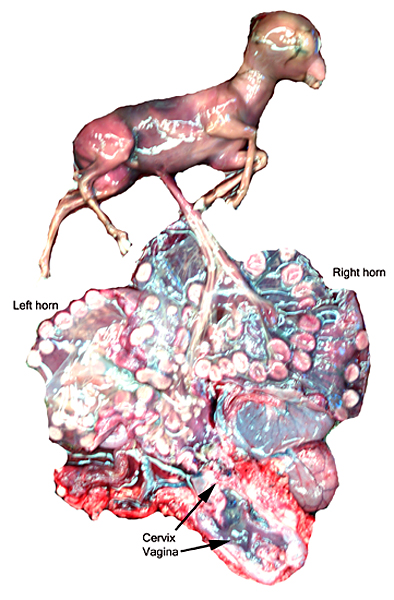
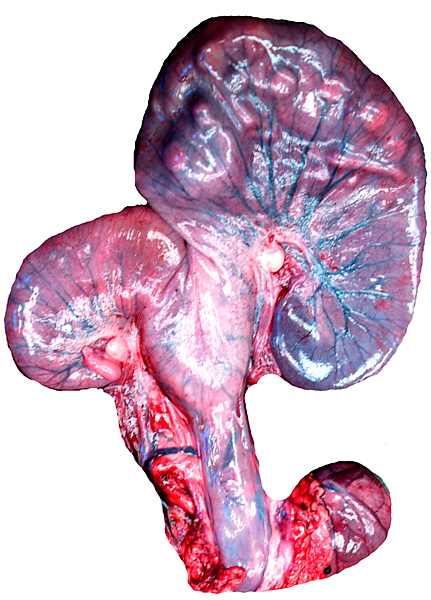
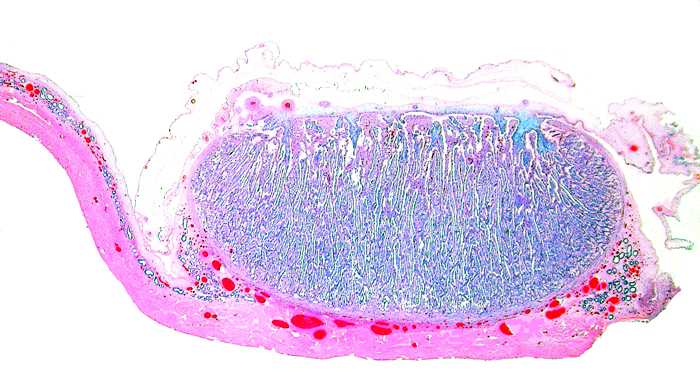
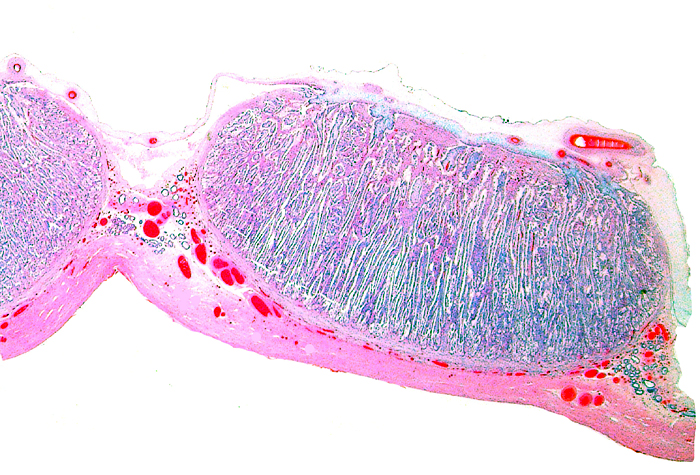
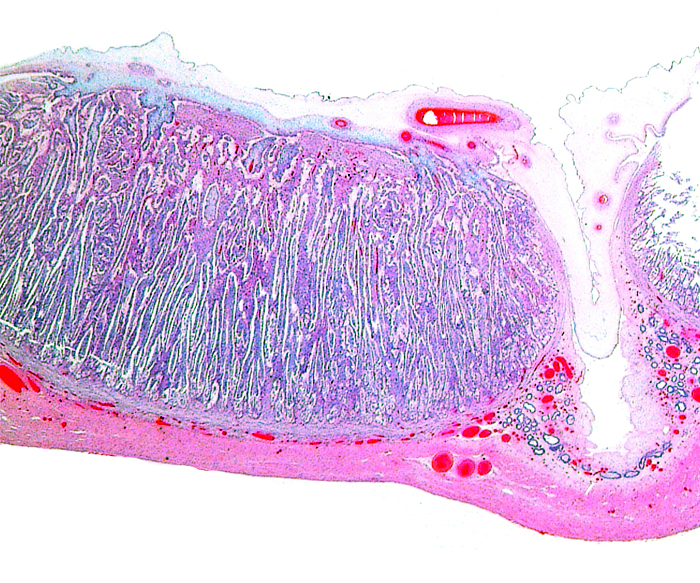
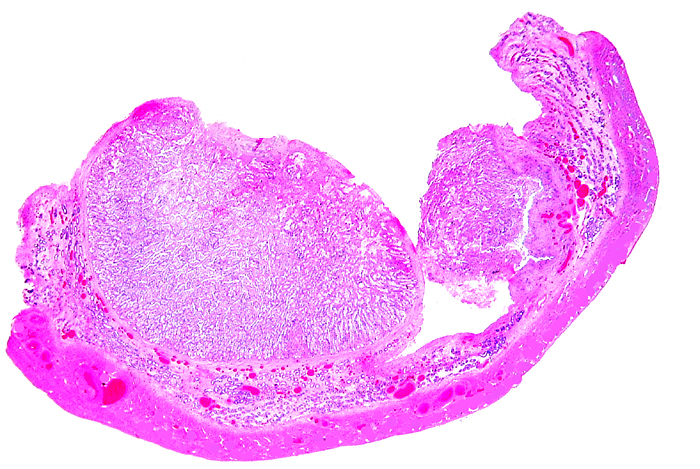
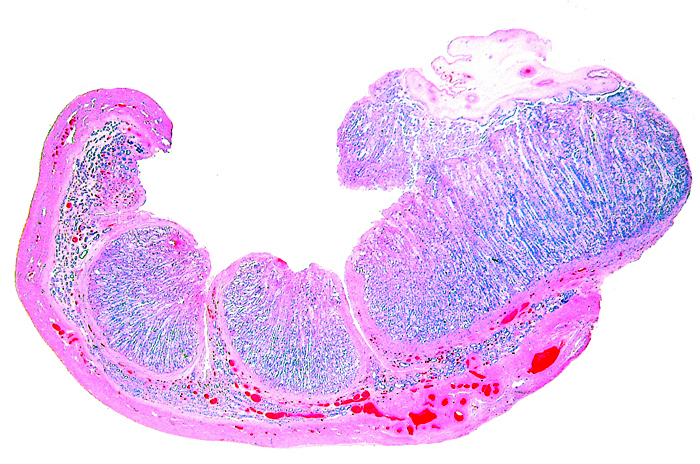
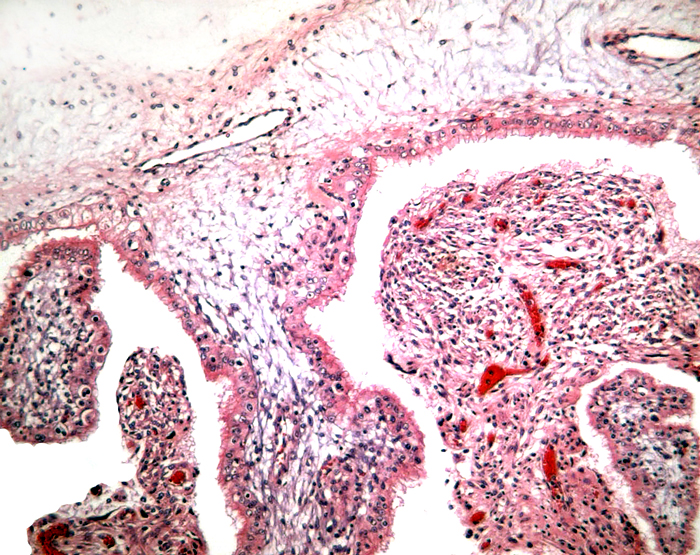
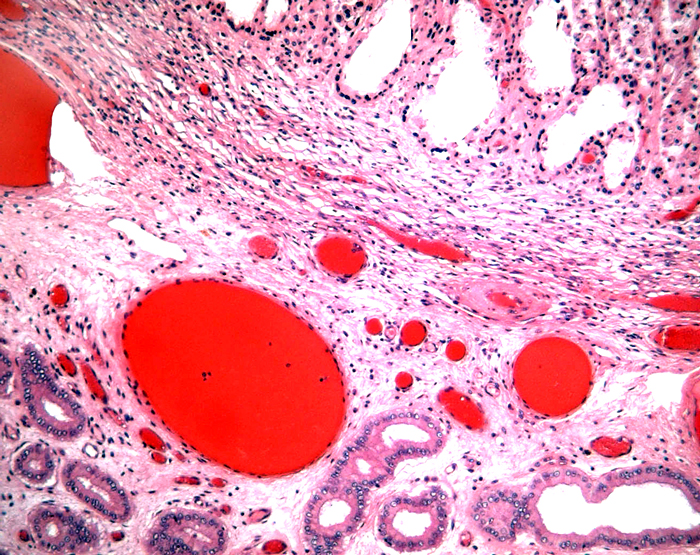
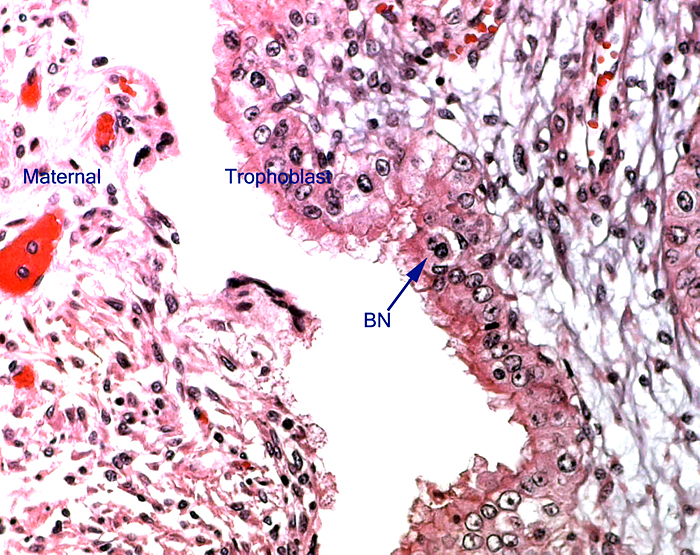
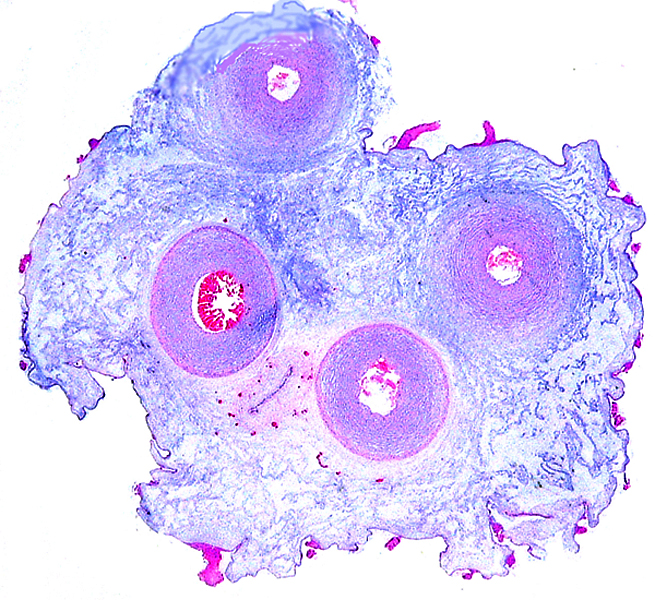
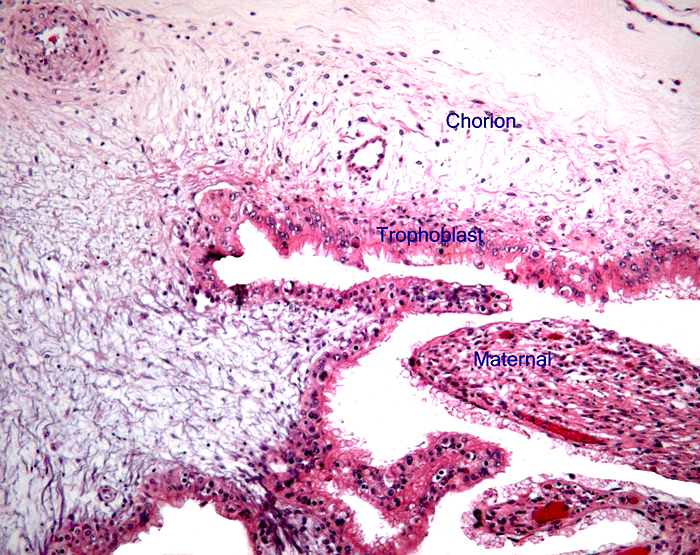
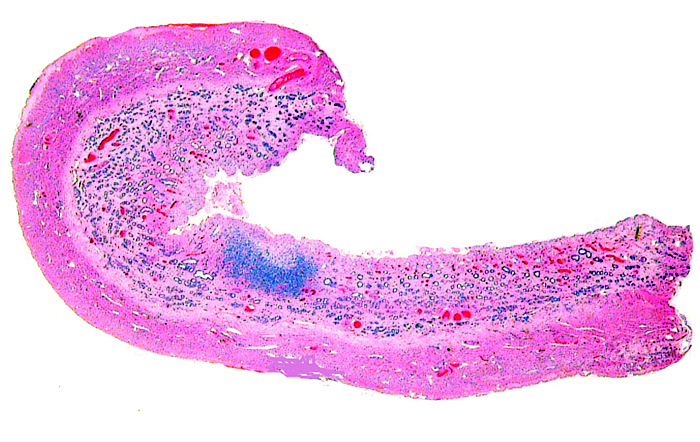
The remarkable feature of the second placenta described here is that, despite the fetus' significant maceration, the placental attachment had not suffered and there was very little degeneration of the villous tissue. Apparently, the endometrial perfusion of the caruncle was sufficient to maintain adequate structure, despite the absence of fetal circulation. The cord and its endothelia and allantoic epithelium had autolyzed.
16) Physiologic data
Pospisil et al. (1984) compiled various hematological parameters of Grant's gazelles and other species.
17) Other resources
Cell strains are available from CRES at the San Diego Zoological Society. We maintain many cell strains of Grant's gazelles at present. They can be obtained by contacting Dr. O. Ryder at oryder@ucsd.edu.
18) What additional information is needed?
Implantational stages and term placentas and their dimensions are unreported so far.
Acknowledgement
The animal photograph in this chapter comes from the Zoological Society of San Diego. I appreciate also very much the help of the pathologists at the San Diego Zoo.
References
Appleby, E.C., Gibbons, L.M. and Khalil, L.F.: Cooperioides antidorca from intestinal nodules in Gazella thomsoni and G. granti . J. Comp. Pathol. 107:295-303, 1992.
Arctander, P., Kat, P.W., Aman, R.A. and Siegismund, H.R.: Extreme genetic differences among populations of Gazella granti , Grant's gazelle in Kenya . Heredity 76:465-475, 1996.
Atkinson, Y.H., Gogolin-Ewens, K.J., Hounsell, E.F., Davies, M.J., Brandon , M.R. and Seamark, R.F.: Characterization of placentation-specific binucleate cell glycoproteins possessing a novel carbohydrate. J. Biol. Chem. 268:26679-26685, 1993.
Baumeister, B.M., Castro, A.E., McGuire-Rodgers, S.J. and Ramsay, E.C.: Detection and control of rotavirus infections in zoo animals. J. Amer. Vet. Med. Assoc. 183:1252-1254, 1983.
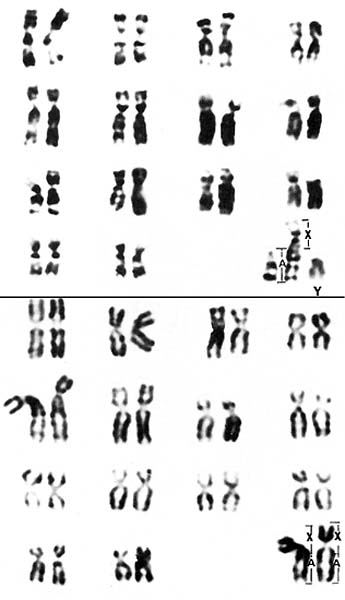
Effron, M., Bogart, M.H., Kumamoto , A.T. and Benirschke, K.: Chromosome studies in the mammalian subfamily Antilopinae . Genetica 46: 419-444, 1976.
Gallagher, D.S. and Womack, J.E.: Chromosome conservation in the Bovidae. J. Hered. 83:287-298, 1992.
Griner, L.A. : Pathology of Zoo Animals. Zoological Society of San Diego , San Diego , California , 1983.
Hayssen, V., van Tienhoven, A. and van Tienhoven, A.: Asdell's Patterns of Mammalian Reproduction: a Compendium of Species-specific Data. Comstock/Cornell University Press, Ithaca , 1993.
Hsu, T.C. and Benirschke, K.: An Atlas of Mammalian Chromosomes. Vol. 9, Folio 439, 1975. Springer-Verlag , New York .
Jones, M.L.: Longevity of ungulates in captivity. Intern. Zoo Yearbk. 32:159-169, 1993.
Lee, S.Y., Mossman, H.W., Mossman, A.S. and del Pinto, G.: Evidence of specific nidation site in ruminants. Amer. J. Anat. 150:631-640, 1977.
Matthee , C.A. and Robinson, T.J.: Cytochrome b phylogeny of the family bovidae: resolution within the alcelaphini, antilopini, neotragini, and tragelaphini. Mol. Phylogenet. Evol. 12:31-46, 1999.
Matthee , C.A. and Davis , S.K.: Molecular insights into the evolution of the family Bovidae: A nuclear DNA perspective. Mol. Biol. Evol. 18:1220-1230, 2001.
Mossman, H.W.: Vertebrate Fetal Membranes. MacMillan, Houndmills, 1987.
Nowak, R.M.: Walker 's Mammals of the World. 6 th ed. The Johns Hopkins Press, Baltimore, 1999.
Pospisil, J., Kase, F., Vahala, J. and Mouchova, I. : Basic haematological values in antelopes – III. The Reduncinae and the Antelopinae. Comp. Biochem. Physiol. A 78:809-813, 1984.
Wilson , D.E. and Reeder, D.A.M.: Mammal Species of the World. A Taxonomic and Geographic Reference. 2 nd ed. Smithsonian Institution Press, Washington , DC , 1992.
Wooding, F.B.: The role of the binucleate cell in ruminant placental structure. J. Reprod.
Fertil. Suppl. 31:31-39, 1982.
Wooding, F.B., Morgan, G. and Adam, C.L.: Structure and function in the ruminant synepitheliochorial placenta: central role of the trophoblast binucleate cell in deer. Microsc. Res. Tech. 38:88-99, 1997.
|